- Research
- Open access
- Published:
Dynamics of a stochastic population model with predation effects in polluted environments
Advances in Difference Equations volume 2021, Article number: 189 (2021)
Abstract
The present paper puts forward and probes a stochastic single-species model with predation effect in a polluted environment. We propose a threshold between extermination and weak persistence of the species and provide sufficient conditions for the stochastic persistence of the species. In addition, we evaluate the growth rates of the solution. Theoretical findings are expounded by some numerical simulations.
1 Introduction
Environmental pollution because of industry, commerce, agriculture, and the rapid growth of human population has been increasingly prominent and has become an global problem. Pollutants have threatened the health of living organisms. For instance, Amoco Cadiz incident carried off more than 260 thousands tonnes of sea animals in one month [1]; Bhopal disaster killed about 15 thousands people and injured about 560 thousands people [2]; according to the United States Fish and Wildlife Service, pesticides carried off more than 72 million birds every year in US [3].
To probe the influence of pollutants on the evolution of populations, many mathematical models have been put forward. In the 1980s, Hallam and his co-workers [4–6] first constructed a series of deterministic models with pollution and uncovered that environmental pollution has serious influence on the persistence and extermination of the species. These results were improved and extended [7–17]. Particularly, motivated by the fact that the evolution of populations often encounters environmental perturbations [18], several authors (see, e.g., [9, 12–17]) paid attention to stochastic population models with pollution and uncovered that environmental perturbations have vital functions on the persistence and extermination of the species.
On the other hand, most populations have parasites and predators [19]. In general, these predation effects have negative functions on the growth of populations. Therefore we need to test population models with predation effects in polluted environments. However, little research has been conducted to exploit this problem (even for deterministic models).
The objective of this paper is testing the above problem. We construct a population model with predation effects in polluted environments in Sect. 2 and testify that the model has a unique global positive solution in Sect. 3. Then we offer a threshold between extermination and weak persistence of the species and provide conditions under which the species is stochastically persistent in Sect. 4. In Sect. 5, we estimate the growth rates of the solution of the model. Finally, we give the conclusions of this paper and numerically expound the theoretical findings in Sect. 6.
2 The model
Without pollution and predation effects, suppose that the growth of the species follows the logistic role:
where \(\Phi (t)\) is the population size at time t, and \(b>0\) and \(\xi >0\) stand for the growth rate and the intraspecific competition rate, respectively.
We consider the predation effects. In general, the predation effects saturate at high prey density and vanish quadratically as the prey density tends to zero [19]. Therefore it is reasonable to model the predation effects by the following function:
where λ is the upper limit of the predation effects, and \(\rho ^{2}\) measures the saturate effect. Then model (1) is replaced by
As said before, the evolution of populations often encounters environmental perturbations [18]. In general, we can take advantage of a color noise process to portray the environmental perturbations [20], and it is suitable to utilize a Gaussian white noise process to depict a weakly correlated color noise [21]. Various ways were developed to incorporate the white noise into deterministic population models. A widely accepted way is to suppose that some parameters in the model are influenced by the white noise (see, e.g., [22–32]). Adopting these approaches,
where \(\psi _{1}(t)\), \(\psi _{2}(t)\), and \(\psi _{3}(t)\) are independent standard Wiener processes defined on a certain complete probability space \((\Omega,\mathcal{F},\{\mathcal{F}_{t}\}_{t\geq 0},\mathrm{P})\), and \(\beta _{i}\), \(i=1,2,3\), stand for the intensities. As a result, model (2) is replaced by
To characterize the influence of pollution, we hypothesize that the populations suck up the pollutants into their bodies [4–6]. Denote by \(T(t)\) the concentration of pollutants in the species, which can lead to a descent of the growth rate [4]:
where \(H_{1}(T)\) is a positive continuous increasing function of T. Accordingly, model (3) is replaced by
To depict the pollutants in the species, we pay attention to the following equation:
where \(T_{e}(t)\) represents the concentration of pollutants in the environment, \(H_{2}(T_{e}(t))>0\) characterizes the suck up of pollutants from the environment, \(H_{3}(T(t))>0\) measures the loss of pollutants because of excretion and detoxication. Both \(H_{2}\in C^{1}\) and \(H_{3}\in C^{1}\) are increasing functions.
Finally, we portray the changes of \(T_{e}(t)\). Denote by \(u(t)\) a continuous and bounded function of t, the input of pollutants from the outside of the environment. Suppose that the changes of \(T_{e}(t)\) are governed by the following equation:
where \(H_{4}\in C^{1}\), measuring the loss of the pollutants from the environment, is an increasing positive function of \(T_{e}\).
According to (4)–(6), we derive the following model:
The objectives of this paper is probing some dynamical properties of \(\Phi (t)\). Note that the last two equations in model (7) do not depend on \(\Phi (t)\), and they have a unique solution \((T(t), T_{e}(t))\) for certain initial value. As a result, from now on we concentrate on Eq. (4).
3 The existence and uniqueness of the solution
Theorem 1
For any initial value \(\Phi (0)>0\), Eq. (4) possesses a unique global positive solution \(\Phi (t)\) almost surely (a.s.).
Proof
We first concentrate on the equation
with \(x(0)=\ln \Phi (0)\). By the locally Lipschitz continuity of the coefficients in Eq. (8) we deduce that Eq. (8) possesses a unique solution on \([0,\tau _{e})\), where \(\tau _{e}\leq +\infty \). It then follows from Itô’s formula (see [33], p. 32, Theorem 6.2) that \(\Phi (t)=e^{x(t)}\) is the unique positive solution of (4).
Now we testify that \(\tau _{e}=+\infty \). Let \(m_{0}\) be a positive constant such that \(m_{0}>\Phi (0)\). For each integer m, define
Let \(\tau _{\infty }=\lim_{m\rightarrow +\infty }\tau _{m}\). We can deduce that \(\tau _{\infty }\leq \tau _{e}\). If \(\tau _{e}<+\infty \), then we can find out constants \(\hat{T}>0\) and \(\epsilon \in (0,1)\) such that
It then follows that we can find out an integer \(m_{1}\geq m_{0}\) such that for any \(m\geq m_{1}\),
where \(\Omega _{m}=\{\omega:\tau _{m}\leq \hat{T}\}\). Define
Then we deduce by Itô’s formula that
Accordingly,
By Gronwall’s inequality (see [33], p. 45, Theorem 8.1) we get
For \(\omega \in \Omega _{m}\), \(W_{1}(\Phi (\tau _{m},\omega ))\geq m^{\gamma }\). Then by (9) and (10) we can see that
Letting \(m\rightarrow +\infty \) gives
However, by the definitions of \(W_{1}(\Phi )\) and T̂ we have
which is a contradiction with (11). Thereby, \(\tau _{e}=+\infty \). □
4 Extinction and persistence
Theorem 2
If \(\Lambda <0\), then \(\lim_{t\rightarrow +\infty } \Phi (t)=0\) a.s., that is, the species is extinct, where
Proof
We can deduce from Itô’s formula that
that is,
where
The quadratic variations of \(\Gamma _{2}(t)\) and \(\Gamma _{3}(t)\) are
and
respectively. We can easily obtain that
By the exponential martingale inequality (see [33], p. 44, Theorem 7.4) we deduce that
By the Borel–Cantelli lemma (see [33], p. 7, Lemma 2.4), for almost all \(\omega \in \Omega \), there exists an integer \(M_{0}=M_{0}(\omega )\) such that for \(M\geq M_{0}\),
Therefore, for any \(0\leq t\leq M, M\geq M_{0}\),
Then we deduce from (12) and (14) that for any \(0\leq t\leq M, M\geq M_{0}\),
For \(0< M-1\leq t\leq M, M\geq M_{0}\), we have
Then (13) shows that
Therefore, for any \(\epsilon \in (0,-\Lambda )\), there is \(T_{1}\) such that for all \(t>T_{1}\),
That is, for all \(t>T_{1}\)
Hence \(\lim_{t\rightarrow +\infty } \Phi (t)=0\) a.s. □
Theorem 3
If \(\Lambda \geq 0\), then
In particular, \(\Lambda =0\Rightarrow \) \(\lim_{t\rightarrow +\infty }t^{-1}\int _{0}^{t} \Phi (s) \,\mathrm{d}s=0\), that is, the species is nonpersistent in the mean.
Proof
For arbitrary \(\epsilon >0\), we can find \(\tilde{T}>0\) such that for \(0<\tilde{T}<M-1\leq t\leq M, M\geq M_{0}\),
Set \(\Xi =\Lambda +\epsilon \). In view of (15), for arbitrary \(0<\tilde{T}<M-1\leq t\leq M, M\geq M_{0}\), we obtain
Set \(\alpha (t)=\int _{0}^{t} \Phi (s)\,\mathrm{d}s\). Then we can see that
Integrating both sides from T̃ to t, we get
Therefore
Taking logarithms leads to
As a result,
It then follows from L’Hospital’s rule that
Since ϵ is arbitrary, we get (16). □
Theorem 4
If \(\Lambda >0\), then \(\limsup_{t\rightarrow +\infty } \Phi (t)>0\), that is, the species is weakly persistent.
Proof
Denote
We hypothesize that \(\mathrm{P}\{L\}>0\). For arbitrary \(\omega \in L\), \(\lim_{t\rightarrow +\infty } \Phi (t,\omega )=0\). Therefore, for any \(\epsilon \in (0,1)\), there is T such that \(\Phi (t,\omega )\leq \epsilon \) for all \(t\geq T\). As a result, for sufficiently large t,
and
where \(M_{1}\) and \(M_{2}\) are positive constants. Consequently,
We then deduce from (12) and (13) that
a contradiction. As a result, \(\mathrm{P}\{L\}=0\).
Denote
and hence \(\widetilde{L}\cap L=\emptyset \). Note that \(\Phi (t)>0\), and therefore \(\widetilde{L}\cup L=\Omega \). Then we deduce from \(\mathrm{P}\{L\}=0\) that \(\mathrm{P}\{\widetilde{L}\}=1\), which is the required statement. □
Theorems 2–4 uncover that Λ is the threshold between extermination and weak persistence of the species. Now let us test the permanence of the species.
Theorem 5
If \(\bar{\Lambda }:=\Lambda _{1}-\limsup_{t\rightarrow +\infty }H_{1}(T(t))>0\), then the species is stochastically permanent, that is, for any \(\epsilon \in (0,1)\), we can find two constants \(\sigma _{1}=\sigma _{1}(\epsilon )>0\) and \(\sigma _{2}=\sigma _{2}(\epsilon )>0\) such that
Proof
Set
Then we deduce by Itô’s formula that
Choose a constant \(\varpi \in (0,1)\) that satisfies
Set
Then we deduce by Itô’s formula that for sufficiently large t,
where
Here \(\epsilon <\bar{\Lambda }-\varpi \beta _{1}^{2}\). Choose a constant \(\nu >0\) that satisfies
Set
Then we deduce bymItô’s formula that for sufficiently large t,
where
According to (18),
Thereby
Accordingly,
For any \(\epsilon >0\), set \(\sigma _{1}= (\epsilon /\tilde{\sigma } )^{\frac{1}{2\varpi }}\). By Chebyshev’s inequality (see [33], p. 5),
Thereby
Accordingly,
Now we testify
Set
Then we deduce by Itô’s formula that
where \(\sigma _{3}>0\) is a constant. Accordingly,
By Chebyshev’s inequality we derive (20). □
5 Upper- and lower-growth rates
Theorem 6
For model (4), we have
Proof
We deduce from Itô’s formula that
Accordingly,
where
Set
Thereby
By the exponential martingale inequality (see [33], p. 44, Theorem 7.4), for any \(\kappa >1\) and \(\varrho >0\),
By Borel–Cantelli’s lemma (see [33], p. 7, Lemma 2.4) we deduce that for almost all \(\omega \in \Omega \), there is \(m_{2}\) such that for any \(m\geq m_{2}\),
Accordingly, for \(m\geq m_{2}\) and \(0\leq t\leq \varrho m\),
Then from (22) it follows that for \(m\geq m_{2}\) and \(0\leq t\leq \varrho m\),
where \(\sigma _{4}=\max \{1,-\ln \xi +b-1\}\). As a result, for \(0<\varrho (m-1)\leq t\leq \varrho m\) and \(m\geq m_{2}\), we derive
Note that
Accordingly,
Letting \(\kappa \rightarrow 1\) and \(\varrho \rightarrow 0\), we obtain (21). □
Theorem 6 probes the upper-growth rate of \(\Phi (t)\). Now let us consider the lower-growth rate of \(\Phi (t)\).
Theorem 7
If \(\bar{\Lambda }>0\), then
Proof
We can deduce from (19) that there is a constant \(\sigma _{5}>0\) such that
By Itô’s formula,
where
Choose a positive constant \(\sigma _{6}\) such that
Accordingly,
Choose a positive constant θ such that
Let \(N=1,2,\dots \). Then from (25) we deduce that
By the Burkholder–Davis–Gundy inequality (see [33], p. 40, Theorem 7.3) we have
Moreover,
Then from (27), (28), and (29) it follows that
For any \(\epsilon >0\), Chebyshev’s inequality suggests that
By the Borel–Cantelli lemma, for almost all \(\omega \in \Omega \), we can find an integer \(N_{0}\) such that for any \(N\geq N_{0}\) and \((N-1)\theta \leq t\leq N\theta \),
Accordingly,
Letting \(\epsilon \rightarrow 0\) results in
Thereby
which is (23). □
6 Conclusions and simulations
In this paper, we have constructed a stochastic single-species model with predator effects in polluted environments. We have probed some dynamical properties of the model, including the existence and uniqueness of the solution (Theorem 1), the threshold between extermination and persistence (Theorems 2–4), stochastic permanence (Theorem 5), and upper- and lower-growth rates (Theorems 6 and 7). To our best knowledge, this paper is the first one to probe population models with predation effect in a polluted environment.
Now let us numerically expound the theoretical findings by the Milstein method [34]. We choose \(H(T(t))=b_{1}T(t)\) and pay attention to the following discretization equation of model (4):
where \(\xi _{1k}\), \(\xi _{2k}\), and \(\xi _{3k}\) are standard Gaussian random variables. We set \(b=0.4\), \(b_{1}=1\), \(T(t)=0.25+0.1\sin t\), \(\xi =0.3\), \(\lambda =0.2\), \(\rho =0.1\), \(\beta _{2}^{2}=0.1\), \(\beta _{3}^{2}=0.2\). For different \(\beta _{1}\), we plot the trajectories of \(\Phi (t)\) in Fig. 1(a)–(d).
-
1.
In Fig. 1(a), \(\beta _{1}^{2}=0.36\), and hence
$$ \Lambda =b-\frac{1}{2} \beta _{1}^{2}-\liminf _{t\rightarrow + \infty }t^{-1} \int _{0}^{t} H_{1}\bigl(T(s)\bigr)\, \mathrm{d}s=-0.03. $$Then from Theorem 2 we deduce that \(\lim_{t\rightarrow +\infty }\Phi (t)=0\). See Fig. 1(a).
Figure 1 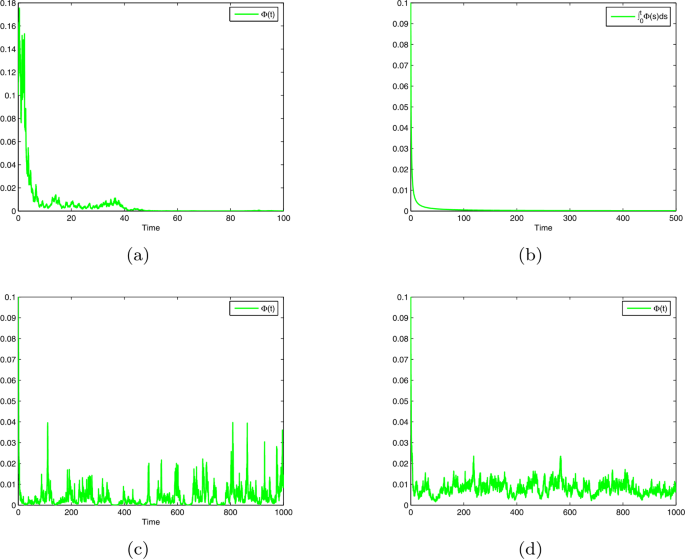
Trajectories of model (4) with \(b=0.4\), \(b_{1}=0.2\), \(T(t)=0.25+0.1\sin t\), \(\xi =0.3\), \(\lambda =0.2\), \(\rho =0.1\), \(\beta _{2}^{2}=0.1\), \(\beta _{3}^{2}=0.2\). (a) \(\beta _{1}^{2}=0.36\), which reflects the extermination of the species. (b) \(\beta _{1}^{2}=0.3\), which reflects the nonpersistence in mean of the species. (c) \(\beta _{1}^{2}=0.25\), which reflects the weak persistence of the species. (d) \(\beta _{1}^{2}=0.04\), which reflects the permanence of the species
-
2.
In Fig. 1(b), \(\beta _{1}^{2}=0.3\), and hence \(\Lambda =0\). Then from Theorem 3 we deduce that \(\lim_{t\rightarrow +\infty }t^{-1}\int _{0}^{t} \Phi (s) \,\mathrm{d}s=0\). See Fig. 1(b).
-
3.
In Fig. 1(c), \(\beta _{1}^{2}=0.25\), and hence \(\Lambda =0.025\). Then from Theorem 4 we deduce that \(\limsup_{t\rightarrow +\infty }\Phi (t)>0\). See Fig. 1(c).
-
4.
In Fig. 1(d), \(\beta _{1}^{2}=0.04\), and hence
$$ \overline{\Lambda }=b-\frac{1}{2} \beta _{1}^{2}- \limsup_{t \rightarrow +\infty }H_{1}\bigl(T(t)\bigr)=0.03. $$Then from Theorem 5 we deduce that \(\Phi (t)\) is permanent. See Fig. 1(d).

To finish this paper, we want to point out that our model (7) may be used to describe the effect of pollution in the usual situation, but it may not well describe some extreme pollution cases (for instance, some extreme examples given in Sect. 1). For these extreme cases, more complicate models should be constructed. We leave these problems for further research.
Availability of data and materials
All data are included in this paper.
References
Maurin, C.: Accidental Oil Spills: Biological and Ecological Consequences of Accidents in French Waters on Commercially Exploitable Living Marine Resources. Wiley, Chichester (1984)
Ichimura, Y., Baker, D.: Acute inhalational injury. In: Reference Module in Biomedical Sciences (2019). https://doi.org/10.1016/B978-0-12-801238-3.11495-3
http://www.fws.gov/birds/bird-enthusiasts/threats-to-birds.php
Hallam, T., Clark, C., Lassider, R.: Effects of toxicant on population: a qualitative approach I. Equilibrium environmental exposure. Ecol. Model. 8, 291–304 (1983)
Hallam, T., Clar, C., Jordan, G.: Effects of toxicant on population: a qualitative approach II. First order kinetics. J. Math. Biol. 109, 411–429 (1983)
Hallam, T., Deluna, J.: Effects of toxicant on populations: a qualitative approach III. Environmental and food chain pathways. J. Theor. Biol. 109, 411–429 (1984)
Freedman, H., Shukla, J.: Models for the effect of toxicant in single-species and predator–prey systems. J. Math. Biol. 30, 15–30 (1991)
Liu, H., Ma, Z.: The threshold of survival for system of two species in a polluted environment. J. Math. Biol. 30, 49–51 (1991)
Gard, T.: Stochastic models for toxicant-stressed populations. Bull. Math. Biol. 54, 827–837 (1992)
Liu, B., Chen, L., Zhang, Y.: The effects of impulsive toxicant input on a population in a polluted environment. J. Biol. Syst. 11, 265–274 (2003)
He, J., Wang, K.: The survival analysis for a population in a polluted environment. Nonlinear Anal., Real World Appl. 10, 1555–1571 (2009)
Liu, M., Wang, K.: Survival analysis of stochastic single-species population models in polluted environments. Ecol. Model. 220, 1347–1357 (2009)
Liu, M., Wang, K.: Persistence and extinction of a stochastic single-specie model under regime switching in a polluted environment. J. Theor. Biol. 264, 934–944 (2010)
Liu, M., Wang, K., Wu, Q.: Survival analysis of stochastic competitive models in a polluted environment and stochastic competitive exclusion principle. Bull. Math. Biol. 73, 1969–2012 (2011)
Zhang, S., Tan, D.: Dynamics of a stochastic predator–prey system in a polluted environment with pulse toxicant input and impulsive perturbations. Appl. Math. Model. 39, 6319–6331 (2015)
Liu, M.: Survival analysis of a cooperation system with random perturbations in a polluted environment. Nonlinear Anal. Hybrid Syst. 18, 100–116 (2015)
Liu, M., Du, C., Deng, M.: Persistence and extinction of a modified Leslie–Gower Holling-type II stochastic predator–prey model with impulsive toxicant input in polluted environments. Nonlinear Anal. Hybrid Syst. 27, 177–190 (2018)
May, R.: Stability and Complexity in Model Ecosystems. Princeton University Press, NJ (2001)
Ludwig, D., Jones, D., Holling, C.: Qualitative analysis of insect outbreak systems: the spruce budworm and forest. J. Anim. Ecol. 47, 315–332 (1978)
Braumann, C.A.: Variable effort harvesting models in random environments: generalization to density-dependent noise intensities. Math. Biosci. 177–178, 229–245 (2002)
Øksendal, B.: Stochastic Differential Equations: An Introduction with Applications, 5th edn. Springer, Berlin (1998)
Beddington, J., May, R.: Harvesting natural populations in a randomly fluctuating environment. Science 197, 463–465 (1977)
Zhu, C., Yin, G.: On hybrid competitive Lotka–Volterra ecosystems. Nonlinear Anal. 71, e1370–e1379 (2009)
Tan, R., Wang, H., Xiang, H., Liu, Z.: Dynamic analysis of a nonautonomous impulsive single-species system in random environments. Adv. Differ. Equ. 218, 1–17 (2015)
Liu, Z., Guo, S., Tan, R., Liu, M.: Modeling and analysis of a non-autonomous single-species model with impulsive and random perturbations. Appl. Math. Model. 4, 5510–5531 (2016)
Liu, M., Zhu, Y.: Stability of a budworm growth model with random perturbations. Appl. Math. Lett. 79, 13–19 (2018)
Lv, H., Liu, Z., Chen, Y., Chen, J., Xu, D.: Stochastic permanence of two impulsive stochastic delay single species systems incorporating predation term. J. Appl. Math. Comput. 56, 691–713 (2018)
Liu, M., Bai, C.: Optimal harvesting of a stochastic mutualism model with regime-switching. Appl. Math. Comput. 375, 125040 (2020)
Li, D., Liu, M.: Invariant measure of a stochastic food-limited population model with regime switching. Math. Comput. Simul. 178, 16–26 (2020)
Ji, W., Hu, G.: Stability and explicit stationary density of a stochastic single-species model. Appl. Math. Comput. 390, 125593 (2021)
Ji, W., Wang, Z., Hu, G.: Stationary distribution of a stochastic hybrid phytoplankton model with allelopathy. Adv. Differ. Equ. 2020, 632 (2020)
Ji, W., Zhang, Y., Liu, M.: Dynamical bifurcation and explicit stationary density of a stochastic population model with Allee effects. Appl. Math. Lett. 111, 106662 (2021)
Mao, X.: Stochastic Differential Equations and Applications. Horwood Publishing, Chichester (1997)
Higham, D.: An algorithmic introduction to numerical simulation of stochastic differential equations. SIAM Rev. 43, 525–546 (2001)
Acknowledgements
The author thanks the editor and the reviewers for their valuable comments.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
The single author is responsible for the complete manuscript. The author read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The author declares that they have no competing interests.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Song, G. Dynamics of a stochastic population model with predation effects in polluted environments. Adv Differ Equ 2021, 189 (2021). https://doi.org/10.1186/s13662-021-03297-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13662-021-03297-w